사과나무의 생활
. 사과나무의 연간 생활주기(生活周期)
사과나무 각 기관의 발육은 재배지역의 기후, 토양에 따라서 또 같은 지역이라도 품종이라
든가 수령에 따라서 차이는 있지만 매년 같은 생활을 되풀이 하고 있다.
사과나무의 1년은 여러 가지 구분방법이 있지만 어느 것이 가장 적절한가는 확실하지 않고
편의상 생육초기인 ‘발아~개화기’, 1년중 가장 영양상태가 불안정하고 그해의 작황을 결정
하는 ‘결실~조기낙과기’, 1년중 가장 온도가 높고 각 기관의 생육도 왕성한 ‘생장ㆍ비대기’,
1년을 마무리하는 시기인 ‘착색~수확기’, 겨울 준비에 들어가 강추위에는 견딜 수 있는 ‘휴
면기’ 등 5개의 기간으로 구분할 수 있다.
사과(후지)의 연간 생활주기
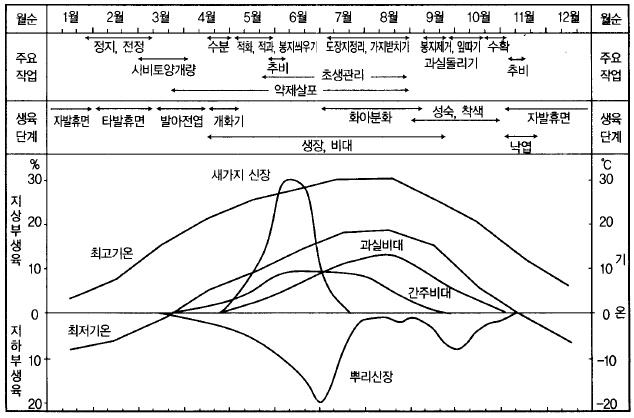
가. 발아~개화기
봄에 춘분(春分)을 지나면서 사과나무의 눈도 급격히 부풀기 시작하여 3월 말부터 4월 상순
에 걸쳐 대부분의 품종이 발아한다. 발아 전에 1회째의 농약살포를 시작하는데 땅속에서는
새로운 뿌리도 신장을 시작한다. 발아 후 10일 정도 지나면 전엽(展葉)이 시작되고, 그 후
4~5일 지나면 벚꽃이 개화하고, 또 10일 정도 지나면 사과꽃이 피기 시작한다.
토양개량이나 기비(基肥)는 전엽기까지 끝내어 비료가 빨리 흡수되어 효과가 나도록 하는
것이 중요하다. 접목은 수액의 흐름이 좋고 유합조직(callus)형성이 활발한 개화 전까지 끝
낸다.
개화가 시작되어 5일 정도 되면 만개기가 되고 또 5일 경과하면 낙화기가 되는데 만개전
2~3일이 인공수분의 최성기이다. 사과나무의 부담을 줄이기 위해서 수분(授分)이 끝나고
수정(受精)이 완료되기 전에 늦게 피는 꽃이나 새가지 끝에 피는 꽃은 화총째 따버린다.
나. 결실~조기낙과기(早期落果期, 6월 낙과)
개화하는 동안 인공수분이나 방화곤충의 활동에 의해서 결실이 된다. 개화나 결실에는 수체
내에 저장되어 있던 영양분이 다량으로 사용되고 또 뿌리의 신장, 새 가지의 비대ㆍ생장,
무수하게 많이 결실된 어린 과실에도 많은 영양분이 소모된다.
한편, 4월에 시용한 비료는 꽤 흡수는 되지만 그것을 이용하여 만들어진 잎은 아직 충분한
영양분을 만들 능력이 없기 때문에 이 시기는 1년중 가장 수체내의 영양이 적은 시기이다.
따라서 사과나무는 쓸데없는 과실을 떨어뜨려(落果시켜) 스스로 영양조절을 하는데 이것을
조기낙과(June drop = 6월낙과)라고 한다.
특히 지난해에 만들어진 저장양분이 부족하든지, 기후가 불순하여 잎에서 만들어진 영양분
이 적든지 하면 심한 조기낙과가 일어나므로 쓸데없는 과실은 일찌감치 적과(摘果)한다.
기온이 높고 수분도 충분하면 지표면의 풀(잡초나 목초)의 생육도 왕성하여 사과나무와 양
수분 쟁탈이 심하여 지기 때문에 풀의 자람을 보아 가면서 풀깍기(刈取)를 한다.
이 시기의 어린과실은 과피가 약하기 때문에 동녹이 발생하기 쉬우므로 사용하는 농약선택
에 주위를 하고 특히 품종 특성상 동녹발생이 많은 품종 (‘감홍’, ‘양광’ 등)은 봉지를 씌워
준다.
다. 생장ㆍ비대 최성기
조기낙과를 피한 과실은 7월부터 9월까지 3개월간 발육이 가장 왕성하며 이 기간 동안 년
간 비대량의 70~80%가 이루어진다. 새가지 신장 및 엽면적 증가는 5월 하순부터 7월 상
순까지 100% 가까이 생장을 완료하고 원줄기 및 주지는 한여름에 비대가 가장 많다. 뿌리
는 6월에 신장이 가장 활발하고 한여름에는 일시적으로 둔화되는데 그때까지 연간 생장량
의 80%가 자란다.
새 가지 신장이 끝나고 7월 상중순부터 화아분화가 시작되면 내년 봄의 개화를 향해서 꽃
눈은 충실해져 간다.
기온이 높은 시기는 광합성, 질소동화, 호흡, 증산, 흡수 등 나무의 생리작용도 매우 왕성하
므로 병해충 방제를 철저히 하여 잎의 건강을 유지한다. 사과잎에 햇볕이 충분히 닿도록 하
여 광합성 능력을 높인다.
잎에 살포한 약액이 잘 묻도록 하기 위하여 방해가 되는 도장지를 정리하고 서로 겹치는 가
지는 지주를 세우든가 유인하여 가지 사이에 공간을 만들어 준다. 이 작업은 과실의 착색에
도 큰 도움을 준다.
한여름에는 수분이 다량으로 소비되어 물부족이 되기 쉬우므로 풀이 지나치게 자라지 않도
록 예취하여 증산을 억제하고, 때로는 관수도 한다.
라. 착색~성숙기
8월 상ㆍ중순부터 조생종의 수확이 시작되는데 이때(음력 7월 보름, 백중)를 경계로 기온은
점차 내려간다. 사과나무나 과실에 저장양분이 축적되어 쓰가루는 8월 중순부터 후지는 9월
상ㆍ중순부터 착색이 시작된다. 잎에서 만들어진 영양분은 과실로 이동하고 저온(低溫), 햇
볕의 힘을 빌려 붉은 색이 만들어진다. 이 시기에는 잎따기 작업이나 봉지벗기기를 하여 과
실착색이 잘 되도록 한다.
과실은 외관 뿐만 아니라, 내용적으로도 경도 저하, 전분 감소, 당도 증가, 산함량의 감소,
식물호르몬의 변화로 낙과가 일어나는 등 점차 과실의 성숙이 진행되는데 성숙한 것부터 수
확을 한다.
한편, 화아는 이 기간 중 더욱 충실해지고 뿌리는 재차 신장을 하지만 과실비대는 점차 떨
어지고 새가지 신장도 물론 정지하고 잎의 기능도 점점 저하하여 엽록소가 분해, 소실해서
황변 낙엽 한다.
중생종의 수확시기에는 아직 잎의 활력이 높지만 만생종의 수확기쯤에는 일부 낙엽이 일어
나고, 일부 기관은 휴면에 들어가 있다.
마. 휴면기(休眠期)
11월에 들어서면 기온도 급격히 내려가고 낙엽도 진행되어 나무는 휴면에 들어간다. 휴면에
들어간 나무는 겨울 추위에 견딜 수 있는 힘이 생기는데 가을이 따뜻한 해에는 휴면에 들어
가는 것이 늦어지기 때문에 갑자기 한파(寒波)가 올 경우에는 어린 가지는 동해를 받아 부
란병으로 연결되므로 과실 수확 후에는 부란병 방제약을 살포하여 준다.
해를 넘겨 월말까지는 자발휴면기 1 (自發休眠期)이지만 따뜻한 날이 나타나는 2월에 들어가
면 나무는 타발휴면기(他發休眠期)에 들어가 눈도 조금씩 부풀어진다.
겨울동안은 저장양분으로 생활을 하지만 이 시기에는 지상부보다 지하부인 뿌리에 양분을
많이 축적하기 때문에 전정에 의한 양분 손실은 적고, 또 나무의 생장에 대한 영향도 여름
전정보다 적다.
타발휴면기에 들어가 내한성이 약하여 졌을 때, 따뜻한 날씨가 지속(暖冬)되다가 한파가 내
습하면 동해를 받기 쉽다. 수체 내에는 눈에 보이지 않는 새로운 움직임이 일어나고 점차
발아를 향해서 눈의 생장이 진행된다.
3. 사과나무의 생장과 영양
‘사과나무는 사과의 나무라야 한다’ 는 의미 깊은 말이 있다. 사과나무는 과실을 생산하는
나무이므로 목재를 생산하는 나무보다도 한층 더 주의를 기울여 관리를 하지 않으면 안된
다.
가. 사과나무의 영양
사과나무가 생활하기 위해서는 인간과 같이 영양분이 필요하다. 태양에너지를 이용하여 잎
이 영양분을 만드는 작용을 광합성이라고 하는데, 이러한 생산공정은 푸른 잎을 가진 식물
이외는 할 수 없는 작용이다.
한편 식물이 광합성으로 생산한 영양 , 즉 당(糖)을 기본으로 하여 단백질을 합성하든지 흡
수, 분해 및 비대 등 여러 가지 활동(일)을 한다. 이러한 활동을 하기 위해서는 호흡에 의해
서 당(糖)을 물(水)과 이산화탄소로 분해할 때 생기는 에너지가 이용된다(그림 11).
광합성은 태양이 나와 있는 낮 동안에 이루어지고, 밤에는 호흡에 의해서 소비만 이루어지
므로 낮에 생산한 량과 호흡 등으로 사용된 량을 뺀 것이 실제로 잎에서 만들어진 물질 생
산량이고, 이것이 식물체의 가장 중요한 영양원이다. 여기에다 토양에 주어진 비료요소 라
든가 원래 토양이 가지고 있던 양분이 흡수되어 생활의 양식(糧食)이 된다.
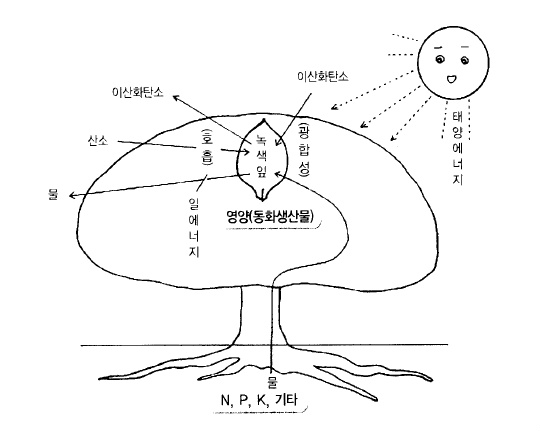
그림 11. 사과나무의 영양분 합성 및 이용의 개략도
나. 영양생장과 생식생장
나무의 생장은 새가지가 뻗고, 수관이 확대되는 영양생장(榮養生長)과 형성되어 꽃이 피고
과실이 맺히는 생식생장(生殖生長)으로 나누어진다. 2가지 생장은 서로 상반되는 면을 가지
고 있는데 영양생장이 너무 강하면 생식생장이 억제되고, 생식생장이 강하면 영양생장이 떨
어진다. 따라서 새가지가 적당하게 자라면서 꽃눈이 붙도록 하는 것이 재배 기술로서 가장
요망되는 것이다.
생장을 조절하는 것은 식물호르몬이라고 알려져 있으나 생장작용의 결과로서 수체 내에는
잎에서 생산된 탄수화물(C)과 뿌리에서 흡수된 질소(N)의 양적관계가 달라지고, 이러한 양
적관계를 C/N율(탄수화물 : 질소 비율)이라고 하며 4가지 형태로 분류한다(그림 12).
Ⅰ형은 질소는 풍부하지만 탄수화물이 부족하여 새가지 생장뿐만 아니라 꽃눈형성, 결실도
불량한 나무이다. 밀실에 의한 일조부족, 수관내부의 가지, 잎이 병해충이나 약해(藥害)를
받은 경우가 이에 해당된다.
Ⅱ 형은 질소는 풍부하지만 질소에 비하여 탄수화물은 약간 부족한 경우로 새가지 생장은
왕성하지만 꽃눈형성이나 결실이 불량한 나무이다. 유목(幼木)과 곧게 선 가지 등이 여기에
해당된다.
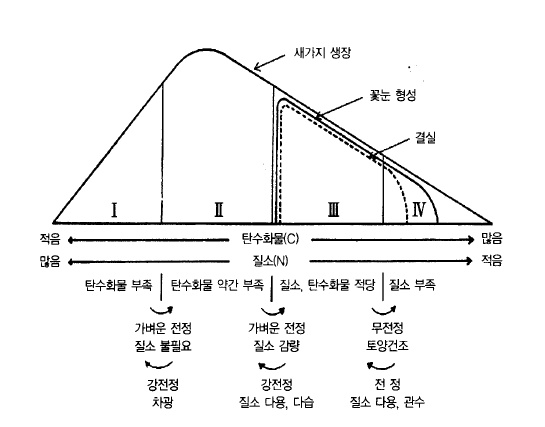
그림 12. 사과나무 체내의 C/N 비율과 인위적 조절법(Gourley, Howlett. 1949)
Ⅲ 형은 질소는 약간 부족하고 탄수화물의 생산은 많은 경우로 새가지 생장은 다소 떨어지
지만 꽃눈형성 및 결실은 극히 양호하여 사과생산에서 가장 바람직한 상태이다. 성목(成木),
수평에 가까이 유인된 성숙한 가지가 여기에 속한다.
Ⅳ 형은 탄수화물은 풍부하지만 질소가 부족하므로 새가지 생장은 떨어지고 꽃눈은 형성되
지만 결실이 불량한 나무이다. 극단적인 상태가 되면 나무는 고사(枯死)한다. 노령수(老齡
樹), 하수지(下垂枝), 흰날개무늬병(문우병)이나 부란병에 걸린 나무, 토양수분의 부족 등 원
인은 여러 가지가 있다.
사과재배에서는 전정, 시비, 방제, 적과, 토양개량 및 기타관리로 항상 Ⅲ 형태의 나무로 개
선, 유지하도록 해야 할 것이다.
다. 저장양분
사과나무는 생육기 뿐만 아니라 휴면 중에도 수체 내에 저장된 양분으로 미약하지만 생활을
계속하고 있다. 이것은 지난해 잎에서 만들어진 영양분이 수체 각 기관에서 쓰고 남은 나머
지가 전분의 형태로 저장되었다가 사용되고 있다.
저장양분은 단순히 생활을 하는데만 쓰이는 것이 아니고 겨울철 내한성을 증가시키고, 화아
를 충실하게 하며 발아, 전엽 및 개화를 촉진시키고 결실을 좋게 하며 과실의 초기발육에
크게 공헌한다.
또, 새가지나 뿌리의 신장도 순조롭게 하는 역할을 한다.
1년중 영양적으로 가장 불안정한 6월에는 저장양분도 가장 적어지는 시기이다. 지난해 만들
어진 저장양분이 적으면 조기낙과 발생이 심하다(그림 13). 이와 같이 저장양분은 초기생육
에 절대적인 영향을 미치기 때문에 잎을 보호하기 위한 병해충 방제, 적과, 시비량 및 적엽
등의 관리에 유의할 필요가 있다.
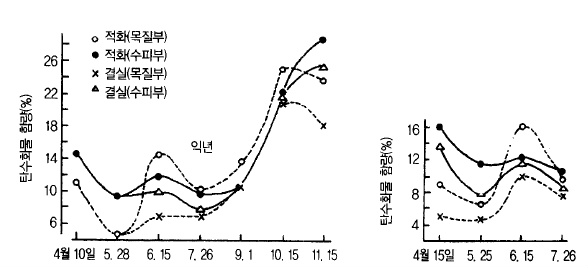
그림 13. 사과나무(홍옥 7년생) 뿌리의 탄수화물 함량의 계절적 변화 (望月1962)
Ⅲ. 계절에 따른 사과나무의 생리ㆍ생태
1. 발아~개화기
가. 발아가 시작된다.
3월 하순경이 되면 사과나무의 눈이 두드러지게 부푼다. 꽃눈은 횡방향으로 부풀어 땅딸막
하게 되고, 잎눈은 홀쭉하여 외관적으로도 서로 구별이 가능하게 된다. 사과눈이 눈에 띄게
부푸는 시기부터 수체내의 수액 흐름도 활발하게 된다. 수액은 뿌리나 가지 속에 저장되어
있던 전분 등이 물에 녹아 당(糖)의 형태로 바뀌어 눈이나 뿌리 선단에까지 운반되어 사과
나무의 생장을 촉진한다.
한편 토양 표층의 뿌리는 의부 온도의 영향을 많이 받아 움직임이 없지만 심층의 뿌리는 3
월 상중순경부터 신장이 시작된다. 이와 같이 외관적으로는 그다지 변화가 없어 보이는 나
무도 눈에 보이지 않는 곳에서부터 활동을 시작하여 3월 말부터 4월 상순에 걸쳐 각 품종
별로 순차적으로 발아를 한다. 발아는 1월 이후 매일의 최고기온에서 6℃를 뺀 수치를 적산
(積算)한 온도와 크게 관련이 있다. 그러나 실제로는 발아기 가까이의 기온일수록 발아에
크게 영향을 미치는 것으로서, 고온일수록 발아가 빨라진다. 특히, 이상고온(異常高溫) 시에
는 예상보다 3~4일 빠른 경우도 드물지 않다.
품종에 따라 발아에 조만(早睌)이 있는데 최근 주요한 재배품종은 발아기가 거의 유사하다.
그러나 같은 지역이라도 수령, 수세, 가지의 성질(단과지, 장과지 등)에 따라 발아기가 다른
데, 유목일수록 발아기는 늦어진다.
눈의 성질에 따라서도 발아기가 달라지는데 액아는 정아보다 1주일 이상 늦게 발아하며, 같
은 정아라도 단과지나 중과지의 정아보다 장과지나 새가지 정아의 발아가 늦어지며, 잎눈은
꽃눈보다 발아가 늦다.
나. 잎이 전개(展葉)된다.
사과잎은 발아하고 나서 10~14일 지나면 전엽이 시작된다. 개화 전까지 6~8매 정도 발생
되는데 이들 잎은 이미 지난해부터 눈 속에 만들어져 있던 것이다. 따라서 엽면적은 작다.
작은 잎은 전엽 약 10일 후, 큰 잎은 약 40일 후에 생육이 완료되는데 양분을 만드는 힘이
약하고, 만든 양분도 대부분 자기 자신의 생장을 위해서 사용되어 버린다.
다. 꽃가루(花粉)가 만들어진다.
꽃눈 속에는 4~6개의 꽃이 포함되어 있다. 수술의 끝에 달려 있는 꽃밥(葯)속에는 화분모
세포가 들어있고, 꽃봉오리가 부풀면서 2개로 분열하는데 핵속에 포함되어 있는 염색체가
반으로 나누어지기 때문에 이러한 분열을 감수분열(減數分裂)이라고 한다. 그 후 다시 한번
분열하여 화분4분자로 되고, 그 4분자가 각각 독립하여 하나의 화분으로 된다. 개화 직전까
지 감수분열이 일어나, 화분의 수가 증가한다.
1개의 꽃밥(葯)속에는 품종에 따라서 차이는 있지만 1만립(万粒) 정도의 꽃가루가 들어있다.
감수분열로 증가한 화분은 꽃봉오리가 붉어질 무렵(pink stage)이 되면 발아 능력을 갖는다
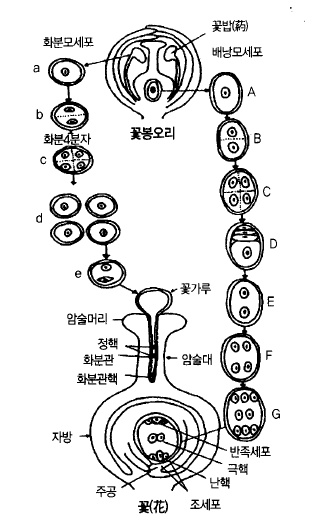
그림 29. 배주(胚珠) 및 화분(花粉)의 완성과 수분까지의 모식도 (熊代, 1977)
한편, 밑씨(胚珠)속에 있는 배낭모세포도 감수분열을 하여 화분과 똑같이 또 한번 분열하여
4개의 세포로 되지만 1개의 세포만 살아남고 나머지는 퇴화된다. 1개의 살아남은 세포는
다시 3회 분열을 하여 8개의 세포로 된다. 1개의 난핵(卵核), 2개의 조세포(助細胞), 2개의
극핵(極核), 3개의 반족세포(反足細胞)로 발달하여 개화 4~5일전에는 수정능력을 가지게 된
다.
라. 사과의 꽃가루(花粉)는 수명이 길다.
사과의 꽃가루는 자연조건에서 1주일 정도 방치하면 50%가, 3주일 정도면 대부분 발아 능
력을 잃어버린다. 그러나 저온ㆍ건조 조건에 저장하면 1년은 충분히 보존할 수 있고 신선
화분과 차이가 없는 발아율을 가진다. 최근에는 화분 저장연구가 진행되어 수년 후에도 변
함없는 발아율을 나타내는 예도 있다. 실용적으로는 0℃이하의 온도, 40~50%이하의 습도
에 저장하면 다음해 수분용의 화분으로서 충분한 역할을 할 수 있다.
마. 꽃가루의 수분(授粉), 발아로 결실된다.
화분은 발아해서 화분관을 신장시킬 만큼의 양분은 스스로 가지고 있다. 화분은 사람 손이
나 곤충, 때로는 바람에 의해서 암술머리에 운반된다. 짚신 모양을 한 화분은 암술머리에
묻으면 수분을 흡수하여 풍선과 같이 부풀어 화분 속에 있던 전분이 분해, 그것을 영양분으
로 발아하여 화분관이 신장한다. 화분관은 암술대를 타고 내려가 밑씨에 도달하면 화분의
내용물이 화분관에 옮겨가서 화분관의 끝에 웅핵(雄核)이 생긴다. 화분관이 밑씨에 닿으면
끝이 터져 웅핵이 밑씨의 난핵과 합체되면 수정이 완료된다(그림 30).
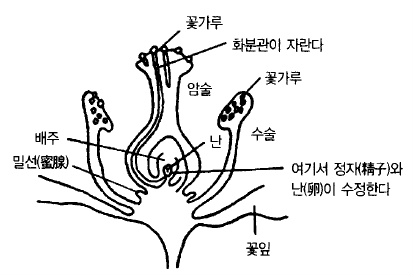
그림 30. 사과 수정(受精)의 모식도(小林)
화분의 발아는 온도가 정도 되면 시작되지만 4~5℃ 화분관이 정상적으로 신장하는 데에는
10℃ 이상의 온도가 필요하고 24~25℃가 최적온도이다. 그러나 28~30℃를 넘어서면 화분
발아는 현저하게 억제된다. 수분(授分)이 된 후 수정(受精)을 완료하기까지의 일수는 대체로
3일 정도 필요하지만 날씨가 좋으면 1일이면 수정이 완료되고, 반대로 날씨가 저온이면 5
일 이상 걸리는 경우도 있다.
밑씨(胚珠)는 개화 4~5일 전 무렵부터 수정능력을 가지며 개화 2~3일전에는 수정능력이
거의 완성된다.
바. 사과는 타가수정이 필요하다.
수정이 이루어지면 씨방(子房)에 배(胚, 종자)가 만들어진다. 그러나 동일품종의 화분이 수
분되면 화분이 발아하지 않든지 발아하여도 화분관이 도중에 신장을 정지하든지 하여 수정
이 어렵게 된다. 이러한 현상을 자가불화합성(自家不和合性)이라고 한다. 따라서 사과에서는
화분관 신장을 순조롭게 하고 수정을 정상적으로 하기 위해서는 다른 품종의 화분을 수분할
필요가 있다(他家受精).
자가불화합성과는 달리 다른 품종의 화분을 수분하여도 수정이 순조롭게 이루어지지 않는
경우가 있다. 이것은 유전적으로 가까운 품종간에 보이는 현상으로 타가불화합성(他家不和
合性) 또는 교배불친화성(交配不親和性)이라고 한다.
그 외에 ‘조나골드’, ‘북두’, ‘하크나인’ 등 3배체 품종은 화분발아력이 극히 불량하여 수분
수로 이용할 수 없다(표 5).

표 5. 사과 품종간 수분(교배) 친화성
주) ◎는 76%이상 결실, ○는 51~75%, △는 26~50%, ×는 25% 이하
※ ◎ 또는 ○은 수분에 적합
사. 곤충은 꽃가루의 운반책이다.
과수에는 드물지만 수분, 수정이 이루어지지 않고, 종자가 형성되지 않아도 결실하는 경우
가 있는데 이것을 단위결과(單爲結果)라고 한다. 온주밀감, 무화과, 감의 평핵무 등이 대표
적인 것이지만 지베렐린(GA)이라는 식물호르몬을 이용하면 포도에서는 무핵포도를 만들 수
있고, 사과에서는 수분시키지 않아도 결실된다. 그러나 사과는 과형이 표주박 형태로 되고
과실에 엽록소가 생성되지 않아 상품가치가 없다.
따라서, 자연상태에서는 다른 품종으로부터 화분이 운반되어 수분, 수정이 되는데, 그 화분
의 운반책이 곤충이다. 사과원의 방화곤충은 백 수십 종이 있지만 주로 화분을 운반하고,
수분에 도움을 주는 것은 꿀벌, 머리뿔가위벌, 꽃등애 등이다.
곤충의 활동은 날씨에 좌우되는데, 고온, 맑은 날씨, 약한 바람과 같은 조건에서 활발하다.
꿀벌은 10℃ 이하가 되면 벌집으로 들어가며, 15℃ 이상으로 바람이 없으면 왕성하게 날아
다니기 때문에 수분도 순조롭게 이루어진다.(그림 31).
즉 저온이고 비가 많고 바람이 강한 날씨일 , 때는 곤충들이 거의 활동하지 않기 때문에 궂
은 날씨가 오래 계속되면 결실이 나빠지므로 사람이 대신에 수분작업을 하지 않으면 안 된
다(그림 32).
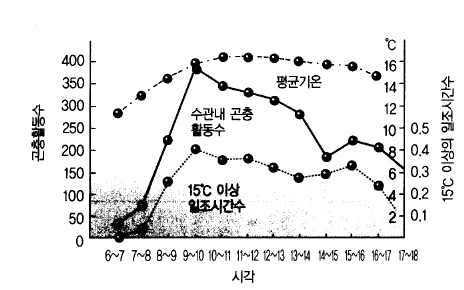
그림 31. 사과원 방화곤충의 비래상황 (일본 아오모리사과시, 1960)
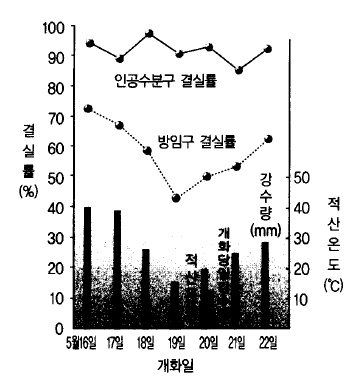
그림 32. 개화기의 기상과 결실률 (일본 아오모리사과시, 1958)
저온이나 다우 에서는 꽃의 (多雨) 생육도 늦어지고, 개약(開葯, 꽃밥이 터지는 것)도 안 된다.
개약 직전의 것은 비가 오면 수분을 흡수하여 갈변하며 속에 들어있던 화분은 발아능력이
떨어지든지, 상실하게 된다. 개약되고 나서 비가 오면 화분이 빗물에 씻겨 흘러내리든지 수
분을 지나치게 흡수하여 파열되므로 결실은 더욱 나빠진다.
2. 결실~조기 낙과기
가. 수정이 되면 꽃받침이 위로 선다.
수정이 되어 종자가 생기고 그 종자에서 식물호르몬이 생성되어 과실세포의 분열을 촉진하
여 드디어 과실비대기로 접어든다. 수정이 이루어진 꽃은 암술머리가 갈변하기 시작하고 1
주일 정도 지나면 꽃받침이 위로 서기 시작한다(그림 33). 수정이 불완전한 꽃은 45도 각도
로 일어설 수는 있으나 완전히 직립하지는 않는다.
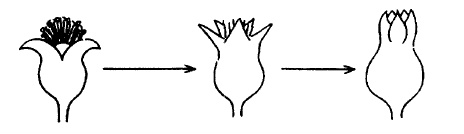
그림 33. 수정이 완료된 과실의 꽃받침 모양 (위로 선다)
나. 새가지가 신장한다.
개화전후부터 신장하기 시작한 새가지는 6월에 들어가면 생장량이 많아지고 대부분의 새가
지는 6월 중순~7월 초순경에 생장을 종료한다(그림 34).
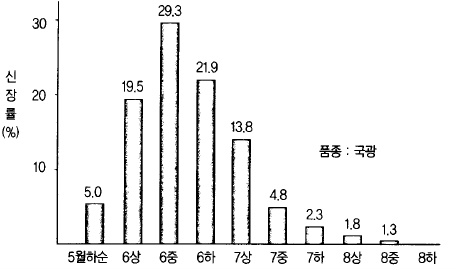
그림 34. 사과나무의 시기별 새가지 신장률 (일본 아오모리사과시, 15년 평균)
수세가 강한 나무나 토양수분이 많은 과수원은 일단 정지한 가지가 재차 신장(2차 신장)을
시작하며, 8월 중하순, 때로는 9월에 들어가서도 신장을 계속한다.
신장을 계속하는 새가지는 약 2.5일마다 1매씩 새로운 잎을 만든다. 잎은 수평 상대(평면)
에서 햇빛을 받기 쉽고 광합성으로 영양분을 생산하기 쉬운 구조로 되어있다. 잎에서 만들
어진 영양분운 일부는 자기 자신이 이용하고 일부는 다른 기관으로 운반되며, 나머지는 저
장양분으로 축적된다.
생육중인 새 가지의 잎은 붙은 위치에 따라 동화양분의 이용장소가 결정된다. 선단으로부터
1/3까지의 잎에서 만들어진 영양분은 새로 생성되는 잎의 생육에 사용되고, 기부의 1/3은
아래쪽 뿌리나 가지의 생장에 이용되며, 중앙의 1/3은 생장정도에 따라 위쪽으로도 아래쪽
으로도 이동한다(그림 36).
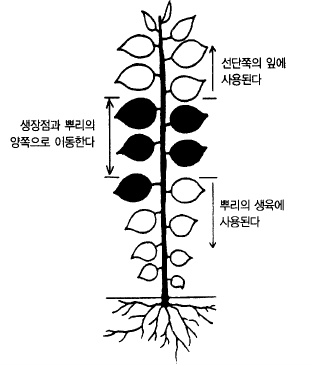
그림 35. 사과나무 잎의 부위별 양분이동 장소
봄에 시비한 비료는 생육이 왕성한 새 가지로. 운반되는데, 인산(P)은 생장점에 모여 가지
의 신장ㆍ비대를 위한 세포분열에 쓰인다. 칼리(K)도 생장점 등 분열이 왕성한 부분에 집중
되어 광합성이나 흡수작용을 높이고, 질소(N)는 식물의 신장ㆍ비대에 큰 역할을 한다.
다. 오옥신(auxin)은 정부우세성(頂部優勢性)을 지배한다.
새 가지의 생장은 정아의 생장이 왕성하고 액아의 생장은 억제되는 정부우세를 나타낸다.
정아의 생장점에서는 식물호르몬의 일종인 오옥신이 만들어져 그것이 가지의 아래쪽으로 내
려가 액아의 생장을 억제한다. 정아를 적심하든지 액아 위쪽에 상처를 주어 오옥신의 이동
을 억제하면 액아가 자라나오는 것을 보아도 그 이유를 이해할 수 있다(그림 36).
그런데 뿌리에서 만들어진 식물호르몬인 시토키닌(cytokinin)을 액아에 뿌려주면 오옥신 때
문에 억제되어 있던 액아가 자라나오기 시작한다. 벤질아데닌(BA, benzyladenin)은 시토키
닌이 포함되어 있는 약제이다.
오옥신이 만들어지는 정아는 오옥신이 다량으로 있는데 어떻게 하여 생장이 계속되는가 의
문이 든다. 그것은 시토키닌이 뿌리에서 이동하여 정아에 많이 모이기 때문일 것으로 생각
된다.
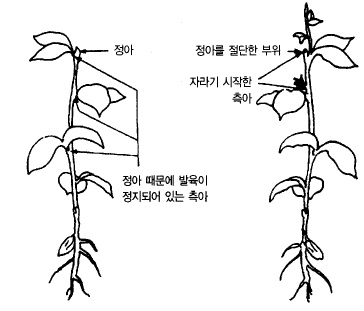
그림 36. 식물의 정부수세성과 타파(打破)
라. 식물호르몬은 작용이 다양하다.
식물호르몬에는 오옥신, 시토키닌 외에 지베렐린, 아브시스산(ABA, abscisic acid), 에틸렌
이 있는데 각각 복잡하게 서로 관련을 가지면서 여러 가지 작용성을 나타낸다.
1) 오옥신(Auxin)
사과 눈(芽)의 생장점에서 만들어져 아래쪽으로 이동하는데 일반적으로는 생장이 왕성한 부
분에 함량이 많으며(그림 37) 발근(發根)이나 캘러스형성에 관여하며 시토키닌과 함께 식물
기관을 만든다.
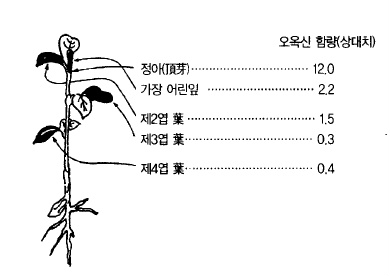
그림 37. 녹색식물의 정아와 잎의 오옥신 분포
잎에 오옥신을 처리하면 이층(離層)형성이 지연되거나 전혀 안되는 등 낙엽을 억제하는 효
과가 있다. 오옥신의 가장 기본적인 작용은 가지의 신장을 촉진하고, 과실의 종자에서 만들
어진 오옥신은 세포의 용적을 확대시켜 과실의 비대를 좋게 한다.
2) 지베렐린(Gibberellin)
벼의 키다리병묘에서 발견되었으며 오옥신과 같이 생장이 왕성한 부위에 함량이 많고 특히
미숙종자에 다량으로 포함되어 있어 오옥신의 신장(伸長)작용을 돕는다. 종류는 70종(1981
년 현재)이상이 있고 종류에 따라서 식물의 신장효과가 상당히 달리 나타나고 있다.
가장 일반적으로 식물에 이용되고 있는 것은 GA3과 GA7이 있고 때로는 GA4도 사용되고 있
다. 지베렐린은 단위결과(單爲結果)를 유발하는 특수한 능력이 있으며 눈이나 종자의 휴면
타파에도 효과가 있다.
사과에는 GA4와 GA7이 들어간 프로말린(주성분 GA4+7, 포미나의 주성분)이 꽃받기 부분의
비대를 촉진시켜 과실을 풍만하게 하는 효과가 인정되고 있다.
3) 시토키닌(Cytokinin)
시토키닌은 미숙종자, 어린뿌리나 잎, 분열조직 등에 포함되어 있는데, 초봄에 포도의 가지
절단면에서 흘러나오는 수액 속에 다량으로 검출되므로 뿌리가 주요한 합성장소로 생각되고
있다.
시토키닌과 오옥신을 함께 처리하면 세포분열이나 비대생장을 촉진한다. 오옥신은 종(縱)
방향으로의 신장을 유도하고 지베렐린은 그 작용을 촉진시킨다. 반면 시토키닌은 오옥신의
작용을 저해하여 횡(橫)방향으로의 생장을 일으킨다(그림 38).
시토키닌의 또 다른 작용은 노화(老化)를 방지한다. 잎은 노화하면 엽록소나 단백질이 분해
되는데 시토키닌은 이러한 노화작용을 방지하여 녹색을 유지하는 특성을 가지고 있다.
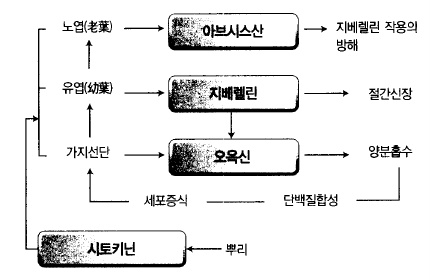
그림 사과나무 38. 새 가지생장을 지배하는 내생호르몬의 상호관계
4) 아브시스산(Abscisic acid)
아브시스산은 앞에서 설명한 3개의 호르몬에 의해서 촉진된 생리현상을 저해하는 작용을
한다. 식물체에 널리 분포하고 있는데 아브시스산 함량은 생장이나 노화 등과 관계가 깊으
며 다른 호르몬과 서로 관련을 가지면서 한다. 예를 들면, 아브시스산에 의한 휴면이나 뿌
리의 생장저해는 지베렐린에 의하여 해제되고 아브시스산에 의한 잎의 노화는 시토키닌에
의해서 억제된다. 이와 같이 아브시스산의 주요한 작용은 식물의 휴면을 일으키고, 이층 형
성을 촉진한다.
최근의 연구로는 아브시스산을 엽면에 살포하면 잎의 증산작용을 억제하는데, 배의 잎에 살
포하면 엽소(葉燒)피해를 방지하는 작용이 인정되고 있다.
왜성대목은 교목성대목보다 뿌리에서 많은 아브시스산이 생성되어 이것이 왜화의 한 요인이
라고 알려져 있다.
5) 에틸렌(Ethylene)
에틸렌은 빛(光)이나 물리적 자극 등으로 생성량이 좌우되는데 상처가 생기면 현저하게 증
가한다. 에틸렌은 식물의 생장을 저해하는 작용을 하며 지베렐린은 에틸렌의 저해작용을 회
복시킨다. 이와 같이, 에틸렌도 다른 식물호르몬과 상호작용을 하는데 특히 오옥신과의 관
계가 깊어 오옥신의 합성이나 이동을 방해한다.
또, 대부분의 식물조직에서 에틸렌이 만들어지는 것은 오옥신의 작용이다. 오옥신이 고농도
로 되면 생장이 저해되는 것은 에틸렌의 생성도 많아지기 때문일 것으로 추정된다.
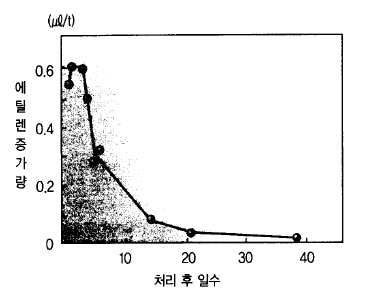
그림 39. 사과나무 가지굽힘과 에틸렌 생성의 관계
(사과나무 가지를 굽히면 그 부위에서 에틸렌발생이 극히 높아짐)
에틸렌은 과실의 성숙이나 호흡상승을 불러일으키고 과숙한 과실에서는 다량으로 만들어 진
다. 저 장중에 과숙과(過熟果)나 상처과(傷處果)가 섞여 있으면 다량의 에틸렌이 생성되어
다른 과실의 성숙을 촉진시킨다. 사과나무의 가지를 굽히면 그 부위에서 다량의 에틸렌이
발생한다는 보고도 있다(그림 39).
마. 기상이 조기낙과의 방아쇠가 된다.
사과의 낙과는 크게 나누어 3시기가 있다. 첫 번째는 수정이 불완전하여 일어나는 불임화
(不稔花) 낙과, 두 번째는 과경이 1~2㎝ 정도일 때 일어나는 조기낙과(6월 낙과, June
drop), 세 번째는 수확 전에 일어나는 후기낙과(수확전낙과)이다(그림 40). 어느 것도 종자
가 적든가, 기능저하로 과실내의 오옥신 생성량이 적기 때문이다.
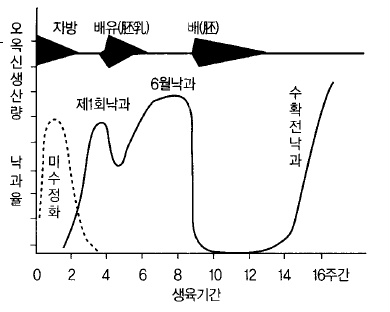
그림 40. 사과에 있어서 오옥신의 생산과 낙과율과의 관계 (Luckwill, 1953)
5월 하순부터 6월 중순에 걸쳐 26~27℃ 이상의 고온이 되고 야간에도 온도가 높아 주야간
(晝夜間)의 온도차가 적은 날이 2~3일 계속되면 과실의 비대가 급격히 떨어지고 그 후
7~10일 경과하면 과경이 황변, 낙과하게 된다.
낙과발생이 많아지는 것은 고온 후에 일조부족이 지속되는 경우에 일어나는데 비가 많이 올
경우 낙과발생이 한층 많아진다. 낙과의 주요인은 기상이지만 과수원의 환경, 개개의 나무
수세, 가지에 따라서 낙과 발생에 차가 있다.
수체내의 저장양분의 부족 잎의 , 동화기능의 저하, 지나치게 강한 수세, 토양의 과습(過濕)
에 의한 뿌리활력 저하 등이 낙과를 조장(助長)하는 요인이다(그림 41).
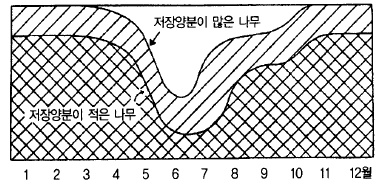
그림 41. 사과나무 수체내의 시기별 저장양분의 소장 (熊代, 1977)
재배적으로는 강전정, 강하게 일어선 가지, 수관내부의 약한 가지, 늘어진 가지(下垂技), 질
소 과잉, 무수분(無授粉), 과잉 착과, 적과지연, 방제 불철저 등도 낙과를 증가시키는 요인
이다. 어쨌든 적정한 수세를 유지하도록 수체관리를 하는 것이 낙과방지의 요점이다. 품종
에 따라서도 조기낙과 발생량이 다른데 ‘축’, ‘세계일’, ‘감홍’ 등이 비교적 많다
바. 동녹이 발생하기 쉬운 시기이다.
낙화기부터 과실의 솜털(毛茸)이 떨어지는 낙화 30일 후까지는 과실에 동녹이 발생되기 쉬
운 시기이다.
1) 사과의 과피(果皮)
과피의 구조는 바깥쪽부터 큐티쿨라, 표피세포(表皮細胞), 하피세포(下皮細胞)로 이루어져
있고, 과피 곳곳에 기공(氣孔)과 솜털(毛茸)이 있다.
큐티쿨라층은 큐틴질이라는 일종의 지방(脂肪)과 같은 물질로 표면에 왁스를 분비하며 물과
공기가 통과하기 어려운 보호조직이다. 낙화 2주 후부터 만들어지는데 층상(層狀)으로 되어
과실이 비대하면서 점차 두꺼워진다. 큐티쿨라층은 기공이나 솜털이 빠진 자리 이외는 틈이
없는 층으로 이루어져 있는데 과실이 비대하면서 균열이 생기며 때로는 표피세포 부분까지
찢어진다(그림 42).
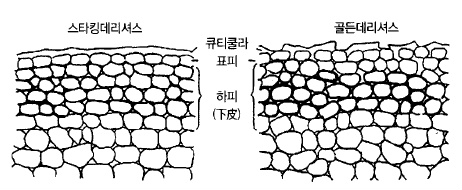
그림 42. 사과 과피(果皮)의 구조(熊代, 1977)
표피세포는 큐티쿨라를 만들며, 사각형 모양의 세포가 1열로 늘어서 있다. 과실이 비대 되
면서 사각형 모양이 횡방향이 길쭉한 곱자(曲尺)모양으로 된다.
과피에 있는 기공은 광합성에 이용되는 이산화탄소 등의 출입은 허용하지만 수분은 필요에
따라서 출입이 조절된다 그러나 점차 기공으로서의 . 기능은 상실되며 일부는 큐티쿨라에 의
해서 기공이 폐쇄되고 대부분의 기공의 가장자리에 코르크 모양의 조직이 생겨 과점(皮目)
이 된다. 특히, 습기가 많은 경우 과점이 커지고 때로는 코르크 모양의 조직이 이상적으로
발달하여 과점 동녹으로 된다.
과피의 솜털(毛茸)은 낙화 3~4주 후에는 대부분 자연적으로 떨어진다. 솜털이 떨어진 자리
는 큐티쿨라층으로 덮이거나 코르크 조직으로 되어 동녹이 생기는 경우도 있지만 보통은 동
녹으로까지는 진전되지 않는다.
2) 동녹의 원인
큐티쿨라층에 어떤 원인으로 상처가 생기면 마치 사람의 상처에 딱지가 생기듯이 외적의 침
입으로부터 조직을 보호하기 위하여 코르크상의 보호조직이 생긴다.
보호조직이 생기는 요인으로는 서리피해, 서로 부딪쳐서 생기는 상처, 전정가위 등에 의한
기계적인 상처, 약해(藥害), 병해(病害), 다우(多雨), 강한 직사광선 등 외적인 것과 과피와
과육조직 발달의 불균형, 유전적인 큐티쿨라조직의 발달 불량 등 내적인 요인이 있다. 실제
동녹 발생은 그러한 요인이 서로 겹쳐 질 때 발생이 많다. 어쨌든 무언가의 원인으로 과피
에 상처가 생겨 과피나 과육세포가 노출되고 그것을 보호하기 위한 조직, 즉 동녹이 생기는
것이다.
동녹은 특히, 과경부에 발생하기 쉬우며 중심과보다 측과에 발생이 많다. 과경부는 과실 비
대가 부위에 따라 차가 생기기 쉽고 측과는 중심과보다 과형의 변화가 크기 때문에 큐티쿨
라에 균열이 생겨 동녹발생이 많다고 생각된다.
3) 동녹 발생시기
골든 데리셔스 의 경우 동녹 ‘ ’ 발생이 많은 시기는 낙화 후 11~30일 간이다. 그 가운데 낙
화 후 16~20일이 가장 동녹 발생이 많다(그림 43). 이 시기는 큐티쿨라층이 불규칙하게 자
라는데 물결모양(波形)이 된다. 또한 표피세포는 직사각형에서 정사각형으로 바뀌어 표피세
포 배열이 흐트러지고 큐티쿨라층과 맞닿는 표피세포는 둥근 모양으로 바뀌므로 큐티쿨라층
에 균열이 생기고 그 보호조직으로서 동녹이 발생한다.
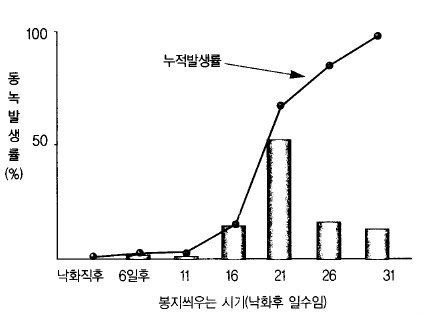
그림 43. 사과 ‘골든데리셔스’의 봉지 씌우는 시기와 과면동녹 발생정도 (일본
아키타과수시, 1973)
사. 잎과 과실의 균형이 중요하다.
6월 말경이 되면 대부분 적과(摘果)가 끝난다. 최근 보급되기 시작한 왜화재배에서는 수세
쇠약이 눈에 띄는 경우가 많은데 원인의 하나로는 과다 착과(過多着果)이다.
잎에서 생산된 동화양분은 각 기관에 분배되어 사용되는데 과실이 지나치게 많이 달리면 한
정된 영양분이 과실에만 많이 이용되어 가지의 생장이 떨어지고 잎이 작아지며 뿌리의 생장
도 억제된다. 물론 과실 간에도 양분 경합이 심해져 과실 하나하나에 대한 양분 공급도 적
어진다.
과실과 잎수 간에 균형이 잡힌 나무는 (葉數) 과실의 무게가 증가할수록 과실의 당함량 특히
비환원당(sucrose)이 증가하고 착색도도 증가한다(표 6).
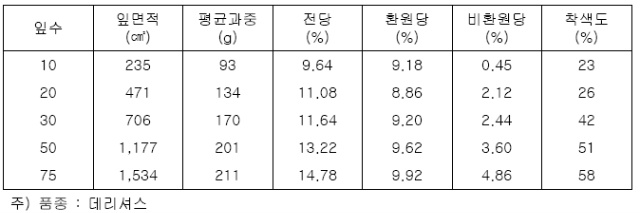
당도가 높은 과실은 초겨울의 한파(寒波)에 의한 동해(凍害)도 적다. 해거리를 방지하기 위
해서도 잎과 과실의 균형은 극히 중요하다. ‘후지’에서는 1과당 잎수가 50매 정도가 되고
그중에서 크기가 30~50㎠인 잎이 20~30% 이상인 것이 바람직하다.
3. 생장ㆍ비대최성기
가. 과실은 급격히 비대한다.
1) 과실의 크기와 모양
사과 과실은 씨방(子房)과 그것을 둘러싸고 있던 꽃받기(花托) 부분이 생장한 것이다. 조생
종의 비대는 직선적인데 비하여 중~만생종은 S자형의 생장곡선을 나타낸다.
과실의 초기생육은 세포분열에 의하여 세포수가 증가함으로써 비대하는데 세포분열은 주로
낮에 이루어지며 만개 3~4주까지 계속된다. 그 후는 분열에 의해서 증가한 세포 하나 하나
가 비대함으로써 과실이 커지고 무게도 증가한다. 세포분열기와는 달리 세포비대는 주로 밤
동안에 이루어지고 낮 동안은 약간 수축된다. 이와 같이 과실의 크기는 세포수와 세포 하나
하나의 크기에 따라 결정되는데 세포용적 확대기부터 과실은 급격히 커진다.
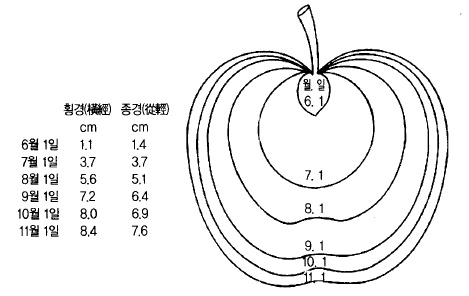
그림 44. 사과 ‘후지’의 시기별 과실비대 정도
과실의 모양도 결실에서 수확까지 상당한 변화가 있는데 세포분열이 끝날 무렵부터 과경이
나 꽃받기 부위에 요철 이 생기고 (凹凸) 지금까지 주로 종축(從軸) 생장을 하던 과실이 횡축
(橫軸)생장을 한다(그림 44, 45).
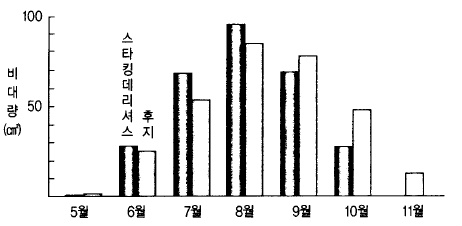
그림 45. 시기별 사과 과실의 비대 (일본 아오모리사과시)
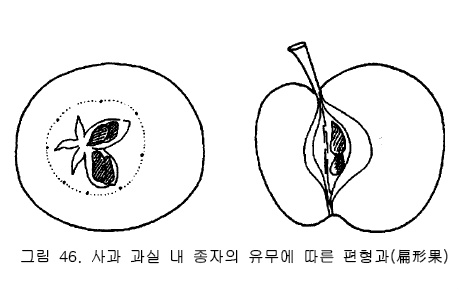
또 사과는 꽃받침 5매, 꽃잎 5매, 수술은 다수 있지만 기본은 5배수, 암술은 하나이지만 끝
이 5개로 나누어져 있으므로 5배수의 꽃이다. 과실도 종자가 들어있는 방이 5개로 나누어
져 있고 각 방에 종자가 들어있는가 그렇지 않는가에 따라 과실의 모양도 바뀐다(그림46).
종자수가 적은 과실은 크기가 작은 과실로 되기 쉬우며, 종자가 들어있지 않은 쪽의 발육이
나빠져 찌그러진 편형과(扁形果)가 된다.
2) 과실비대에 미치는 요인
과실비대의 최적 온도는 20℃이므로 낙화기에서 6월까지와 성숙기 동안은 기온이 다소 높
은 쪽이 좋고, 한여름 동안은 낮은 쪽이 과실 발육에 좋다. 낮 동안 고온이면 세포분열이
활발하고 밤 동안 고온이면 세포의 용적확대가 왕성하여 진다고 한다.
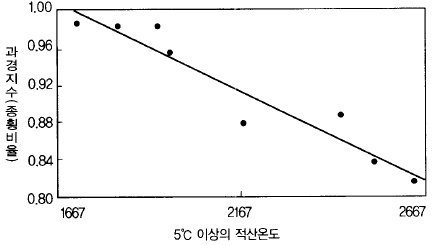
그림 47. 사과 과경(果徑)에 미치는 만개에서 수확까지의 적산온도
(품종 : 데리셔스, westwood 1978)
따뜻한 지방 에서 생산되는 (暖地) 과실은 납작한 과실(扁平果)이 많고 추운 지방(寒冷地)에서
는 종축이 긴 과실이 많은데, 기온이 높으면 횡축 생장이 좋고 기온이 낮으면 종축 생장이
순조롭기 때문이다(그림 47).
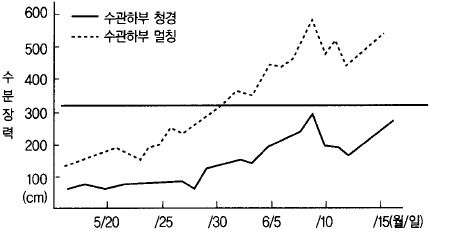
일조가 부족하면 잎의 광합성 능력이 떨어진다. 비 오는 날이 계속되면 일조가 부족함과 동
시에 토양수분이 과다하게 되어 사과나무 뿌리의 활력이 떨어져 양수분의 흡수도 불량하게
되고 과실비대도 나빠진다.
그림 48. 토양수분의 상태(토심 30㎝)와 과실비대
사과나무의 이상은 수분이기 때문에 80% 증산작용으로 잃는 수분은 보급하여 줄 필요가
있다. 따라서 순조로운 생육을 계속하기 위하여 토양 내에 항상 적당한 수분이 유지되어야
한다(그림 48). 가뭄이 지속되면 과실내의 수분은 새 가지의 생활에 사용되므로 과실의 비
대는 극히 떨어진다.
이상과 같이 환경조건은 과실비대에 크게 영향을 미친다. 저장양분이 많고 꽃따기(摘花)를
하고 과실 내 종자가 많으면 과실의 세포수 증가에 큰 영향을 미치며, 적과(摘果)를 조기에
하거나 수관하부에 멀칭을 하면 과실세포의 용적확대를 촉진시킨다.
나. 꽃눈(花芽)은 여름에 만들어진다.
1) 꽃눈형성
새가지가 자라고 잎이 만들어지면 잎자루가 붙은 자리(葉腋)에 눈이 생긴다. 그 눈은 다음
해 정아(頂芽)가 되는데, 수체내의 영양분이 충분하면 꽃눈이 되어 그 다음해에 꽃이 핀다.
사과나무는 눈이 생긴 후 햇수로 3년, 만2년째에 꽃이 핀다. 꽃눈은 어떻게 만들어지는가는
아직 충분히 밝혀져 있지 않지만 잎에서 만들어진 영양분이 어느 정도 축적되면 화성물질
(花成物質, florigen)이 생성되어 양분의 이동 통로인 사관(篩管)을 통하여 눈에 운반되어 꽃
눈이 만들어진다고 한다. 과실이 전혀 착과되지 않아도 5~6월 경에 우박 등으로 인하여 잎
의 기능이 극단적으로 떨어지면 꽃이 만들어지지 않는 것을 보아도 꽃눈을 만드는 데에는
건전한 잎이 필요하다는 것을 알 수 있다. 바꾸어 말하면 꽃눈 형성은 잎의 지령(指令)에
의하여 움직이고 있다고 하여도 좋다. 양분축적과 함께 최근에는 식물호르몬도 화아형성에
밀접한 관계가 있다고 알려져 있다. 정아가 꽃눈으로 되는 데에는 새 가지 생장이 일찍 멈
추고 잎(若葉)에서 만들어지고 있는 지베렐린의 합성이 정지되고 뿌리에서 생산되고 있는
시토키닌이 가지의 수액 내에 남아 있을 필요가 있다(그림 49).
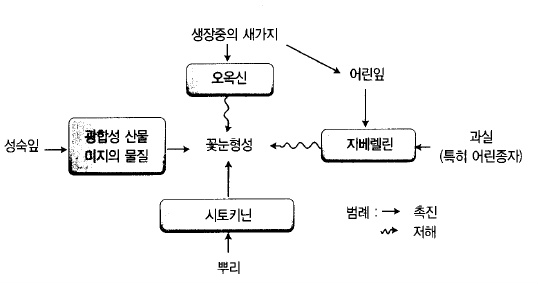
그림 49. 꽃눈 형성을 지배하는 내적요인
또, 단과지 상의 정아는 그 단과지에 과실이 달려 있으면 과실의 종자에서 생성된 지베렐린
이 꽃눈형성을 억제한다. 따라서 가능하면 종자가 생기기 전, 꽃이 피어있을 동안에 적화
(摘花)를 하면 꽃눈이 많이 만들어지게 된다.
2) 꽃눈 분화시기
꽃눈 분화 시기는 품종에 따라 다소 다르지만 7월부터 시작된다. 동일한 품종에서는 따뜻한
지방이 추운 지방보다 빠르고, 고온, 건조한 해일수록 빠르다. 같은 나무 중에서도 단과지,
중과지, 장과지 정아, 액아의 순으로 분화한다.
1개의 눈에서는 먼저 중심화가 분화되고 제1측화, 제2측화의 순으로 되며 가장 늦은 것은
9월에 들어가서 분화한다. 1개의 꽃 중에서는 꽃받침, 꽃잎, 수술, 암술의 순으로 바깥쪽 기
관의 원기부터 만들어지면 꽃밥 내의 꽃가루나 씨방내의 배낭은 꽃이 피기 전에 만들어진
다.
3) 꽃눈의 분화량
기상적으로는 고온, 단일, 일조부족에서는 꽃눈형성이 억제되고 반면 극단적으로 가물지 않
는 한 건조상태가 꽃눈의 수를 많게 하고 7월 중순의 최저기온이 낮을수록 분화량은 증가
한다.
재배적으로는 질소분이 과다하든지 극단적으로 적든지 하면 꽃눈형성이 억제되며 같은 질소
라도 질산태쪽이 암모니아태보다 화아량이 적게 된다고 하는 보고도 있다.
한편, 왜성대목, 가지의 수평유인, 새가지 비틀기(捻枝), 스코어링(환상절피), 환상박피, 아상
(눈 밑에 하는 것)(그림 50), B-9 살포 등은 화아를 증가 시키는 기술이다.
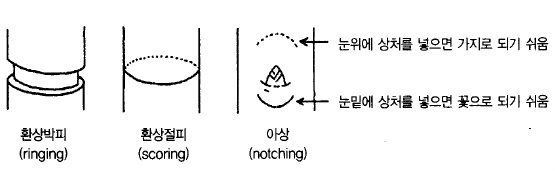
그림 50. 사과의 꽃눈을 증가시키는 처리
4) C/N율
영양생장이나 생식생장을 조절하는 것은 식물호르몬이기 때문에 재배적으로 적절하게 호르
몬이 생성되도록 조절하는 관리를 고려하여야 된다. 식물호르몬에 의한 생장조절의 결과로
서 사과나무의 수체는 다양한 변화를 하는데 잎에서 생산된 탄수화물(C)과 뿌리에서 흡수된
질소(N)의 양(量)적인 비율로 분류한 것이 C/N율이다.
이상적인 나무는 나무의 생장은 다소 떨어지지만 꽃눈의 착생이 좋고 개화, 결실이 양호한
수세를 유지하는 것이다. 수세가 지나치게 강하든지 쇠약할 경우 전정, 시비, 방제, 토양관
리, 착과 조절 등 나무의 상태에 따라서 적절한 관리를 하여 항상 이상적인 나무가 유지되
도록 하는 것이 필요하다.
다. 사과나무의 가지와 줄기가 굵어진다.
가지와 줄기는 잎과 열매를 달고 수분이나 영양분의 통로가 되는 도관(導管)과 사관(篩管)이
있는 부위이다. 도관이나 사관의 세포막이 두꺼워지고 단단하게 되어 식물의 조직을 튼튼하
게 한다.
사과나무의 가지와 줄기는 형성층의 분열에 의하여 비대하는데 기온이 낮으면 분열력이 약
하고, 높아지면 분열하는 힘도 강하여지는데 여름에 분열한 세포는 크고, 추워지면서 작아
진다. 겨울동안에는 거의 세포가 증가하지 않는다. 이와 같은 생장이 매년 되풀이 되므로
겨울동안 생장하지 않는 부분이 경계선이 되어 연륜(年輪, 나이테)으로 나타난다.
10년생 이상된 가지나 줄기의 목부(木部)는 바깥쪽에서 헤아려 9~10년 정도까지의 부분은
양수분을 수송하는 힘이 강하나 그 안쪽으로 갈수록 약하여 진다. 이 부분을 심재(心材)라
고 하며 나무를 튼튼하게 지지하는 역할을 한다. 나무 껍질부분(皮部)은 안쪽으로 갈수록
젊은 부분이고 바깥쪽의 오래된 부분은 점차 조피가 되어 탈락한다. 조피(組皮) 틈은 해충
의 서식 장소가 되므로 봄에 인위적으로 긁어준다.
가지나 줄기도 잎에서 만들어진 양분으로 생활하기 때문에 잎이 많이 붙어 있는 가지, 결실
량이 적은 가지의 비대가 좋고 새로 나온 가지는 뿌리에 가까운 것, 등쪽(背面)에 발생한
가지의 비대가 좋다.
라. 사과나무는 여름에 많은 량의 물을 소비한다.
여름이 되면 증산작용에 의하여 잎에서 많은 량의 물이 소실되며 뿌리가 토양중의 수분을
흡수하여 부족분을 보급한다. 증산작용의 주역인 잎은 7~8월경에 생장이 완료되며 이때가
1년 중 최대의 엽면적을 나타낸다(그림 51).
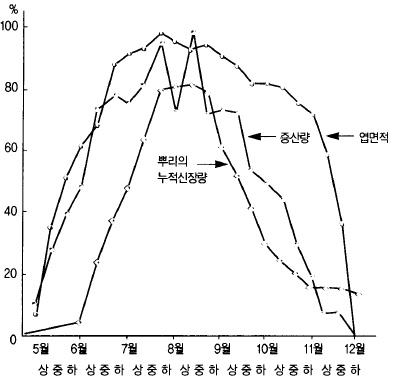
그림 사과나무의 51. 증산량과 시기별 엽면적 및 뿌리의 신장량
(일본 아오모리사과시, 1952)
10a당 70~90만 매의 잎에서 8월 한 달 동안에 약 40㎜(10a당 40톤의 물을 준 것과 같은
양)와 같은 양의 물이 증산작용에 의하여 공기 중에 방출된다. 한편, 강수량의 약 20% 정
도가 식물의 증산작용에 이용된다고 한다.
우리나라 사과주산지의 8월 강수량은 대체로 200~300㎜로 강수량의 20% 정도인 40~60㎜
로 증산량은 충족시킬 수 있으나 최근 국지성 집중호우가 많아 단시간 내에 많은 비가 오기
때문에 8월에도 자연강우에 의한 물 부족 해결은 기대하기 어렵다. 따라서 여름철에는 가능
한 한 자연강우 이용을 높여야 하는데 이를 위해서는 토양의 보수력을 높이는 한편 지표에
서 수분증발을 최대한 방지하는 것이 중요하다.
수분의 흡수와 함께 비료성분의 흡수도 왕성하게 되는데 여름철에 비료성분 특히 질소성분
이 다량으로 흡수되면 가지의 2차 신장을 촉진하여 꽃눈형성을 나쁘게 하고 가지가 무성하
여 수관 내부에 햇볕이 잘 들어가지 않고 병해충 발생이 많아진다. 과실의 착색에도 나쁜
영향을 미치기 때문에 질소성분의 흡수를 억제하기 위하여(그림 52) 추비 시기는 6월 상순
까지는 끝내도록 하여야 한다.
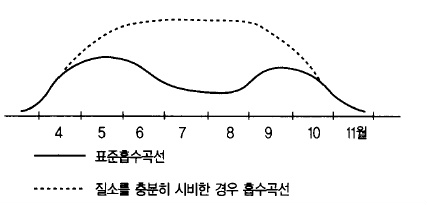
그림 52. 질소의 표준흡수곡선 모식도
4. 착색~성숙기
여름 동안 대부분 녹색으로 덮여 있던 사과원도 가을에 접어들면서부터 생리작용에 큰 질적
인 변화가 일어나며 과실의 외관 및 내용에도 변화가 커진다. 과실의 과육은 부드러워지고
당도가 증가하며 산미가 감소하머 과피색이 붉은 색으로 착색된다.
과실의 성숙은 기상이나 토양, 재배관리 방법에 따라 차가 생기는데 관리의 좋고 나쁨이 외
관이나 식미를 좌우하게 된다.
가. 과실은 숨을 쉰다.(호흡 한다).
사과는 완전히 부패할 때까지 삶을 계속하기 때문에 항상 호흡을 한다. 세포분열이 왕성한
유과기는 호흡이 극히 왕성하지만 과실이 커짐에 따라 호흡량은 급격히 저하한다. 그러나
성숙되기 직전에 재차 호흡이 상승하는데 이러한 호흡상승을 클라이맥터릭 라이즈
(climateric rise, 호흡상승기)라고 한다(그림 53)
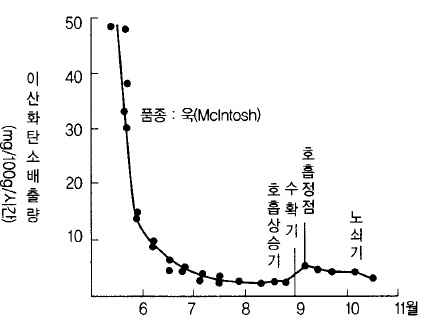
그림 53. 사과 과실의 성숙에 따른 호흡강도의 변화 (Krotkov, 1941)
모든 과실에서 이와 같은 현상을 보이는 것은 아니고 사과 품종 중에서도 조~중생종은 비
교적 확실한 호흡상승 현상을 보이지만 만생종은 그 정도가 낮다.
호흡상승의 원인물질은 에틸렌인데 과실 내에서 호흡상승 직전에 에틸렌 함량이 높아진다.
호흡상승이 최고점에 이르렀을 때를 클라이맥터릭 맥시멈(climateric maximum 호흡정점)이
라고 부르며, 과실이 완숙한 시기와 거의 일치한다.
나. 식물호르몬 함량이 증가하거나 감소한다.
성숙을 일으키는 원인은 식물호르몬이다. 에틸렌 함량이 증가하면 사과에서는 호흡량이 높
아지지만 생장에 관여하는 오옥신, 시토기닌, 지베렐린은 성숙기로 감에 따라 감소한다. 생
장을 저해하는 ABA도 성숙이 진행됨에 따라 감소하지만 수확 전에는 재차 급격히 증가한
다.
따라서 사과의 성숙은 오옥신과 에틸렌, ABA의 양적인 관계로 과실의 성숙이 시작된다.
다. 과실의 내부 성분이 변화한다.
과실의 가장 많은 부분을 차지하는 것은 수분으로서 85% 내외인데 전 생육기를 통하여 그
다지 변화가 없다. 주요한 성분은 탄수화물로 전분, 당, 섬유질, 펙틴질 등이며 그밖에 유기
산도 상당히 포함되어 있는데 당과 산의 비율이 과실의 식미를 좌우한다. 기타 성분으로 단
백질, 비타민, 무기성분 등이 포함되어 있지만 양은 적다. 주요한 성분의 시기적 변화는 다
음과 같다.
1) 전분(澱粉) 함량
전분 함량은 생육초기부터 조금씩 증가하여 한여름에 최고에 달하였다가 점차 감소하여 성
숙과 함께 소실된다(그림 54).
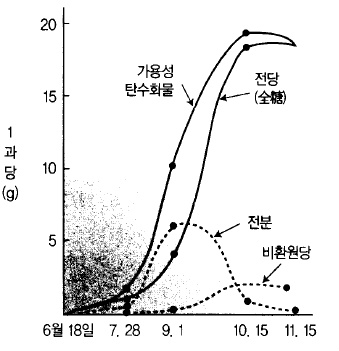
그림 54. 사과 과실내의 탄수화물 함량의 시기별 변화
2) 당(糖) 함량
생육 초기에는 과실내 당 함량이 극히 적지만 성숙이 진행됨에 따라 점차 증가한다. 당 함
량은 광합성으로 만들어진 탄수화물이 전류되기 쉬운 형태(소르비톨, sorbitol)로 바뀌어 과
실에 운반되어 효소의 작용에 의하여 당으로 바뀜으로써 그 함량이 증가한다. 따라서 과실
에 당 함량이 증가되기 위해서는 잎이 없어서는 안 되기 때문에 지나친 적엽(摘葉), 잎따기)
을 하지 않도록 하여야 한다.
사과 과실에 포함된 당의 종류로는 환원당인 포도당(glucose)과 과당(fructose), 비환원당인
자당(sucrose)이 대부분이며 그중 가장 많은 것이 과당인데 수확기까지 그 함량이 증가하며
자당은 성숙이 진행됨에 따라 급격히 증가한다(그림 55).
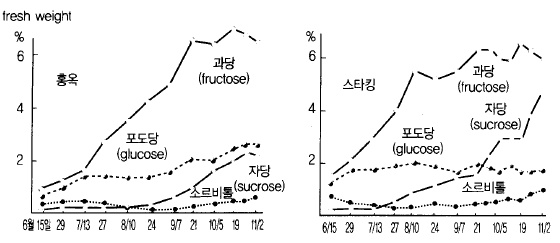
그림 55. 사과 과실 내 당류(糖類)의 시기별 함량
(일본 과수시모리오카지장, 1977)
3) 유기산의 함량
사과에 포함되어 있는 산은 대부분 사과산(malic acid)인데 전분과 같이 여름에서 성숙기에
걸쳐 점차 감소한다. 산의 함량은 품종에 따라 차가 큰데, 조생종은 높아서 1%가 넘는 것
도 있는 반면, 적은 품종은 0.2~0.3%인 것도 있다. 대부분의 품종은 수확 시에 0.3~0.6%
정도이다. 산 함량이 감소하는 것은 과실이 호흡하는데 사용되기 때문이며 고온 조건과 같
이 호흡량이 많은 시기에는 감소폭이 크다. 특히 사과산은 온도에 민감하게 반응하는 산
(酸)이다.
4) 경도의 변화
펙틴(pectin)과 섬유질은 과피에 특히 많으며 과실의 경도를 유지하는 성분이다. 펙틴 화합
물은 과실이 익지 않은 동안은 물에 불용성인 프로토펙틴이 많은데 성숙이 진행되면서 가용
성인 펙틴으로 바뀐다.
펙틴은 과실의 세포를 서로 연결하는 접착제 역할을 하고 있는 화합물이기 때문에 수용성으
로 바뀌면 결합력이 약해져 과실의 경도를 저하시키는 요인이 된다(그림 56).
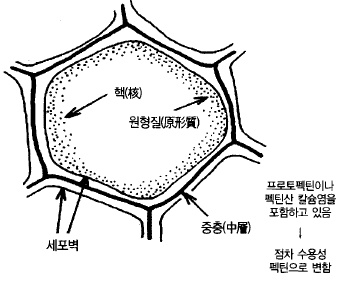
그림 56. 세포의 모식도 (態代, 1977)
5) 비타민C
비타민C는 대부분이 과피 또는 과피 바로 아래의 과육에 포함되어 있고 과실이 자라면서
총 함량은 점차 감소한다. 비타민C의 함량은 환경에 따라 크게 좌우 되는데 그늘에 있는
과실보다 햇볕을 잘 받는 부위의 과실이, 봉지를 씌운 과실보다 씌우지 않은 과실에 많다.
라. 과실에 밀(蜜)이 생긴다.
과실은 성숙되면서 당이 많아진다. 이것은 잎이 태양의 힘을 빌려 만든 광합성산물이 전류
되기 쉬운 소르비톨 형태로 바뀌어 과실로 이동한 다음 효소의 작용에 의해서 당으로 변화
하기 때문이다(그림 57).
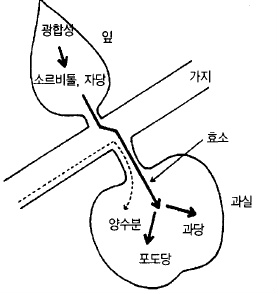
그림 57. 사과나무에 있어서 동화 물질의 전류
맑은 날이 계속되면 잎에 다량의 소르비톨이 생산되어 과실에 축적되는데 효소의 작용이 약
하면 공기로 채워져 있던 과육의 세포간극에 소르비톨이 전류되어 밀증상을 나타낸다.
수확 후 저장 중에 밀은 자연적으로 소실되지만 밀이 다량으로 있을 경우 소실되는 것도 늦
어진다. 따라서 과실세포는 산소를 받아들이기 어렵게 되고 산소결핍을 초래하여 호흡을 할
수 없게 된다.
과실은 살아가기 위해서 세포속에 포함되어 있는 산소를 이용하는 분자간호흡(分子間呼吸)
을 하여 발효(醱酵)가 시작되고 그 결과 알데히드 등 유해물질이 축적되어 과육이 갈변되는
장해로 나타난다.
마. 온도가 낮아지면서 착색이 좋아진다.
사과의 과피색은 적색과 황색이 대부분인데 재배상 어려운 것은 적색 품종이다. 적색 품종
은 여러 가지 요인에 의하여 착색이 좌우된다.
1) 과실의 색소
대부분의 사과 품종은 녹색, 황색 및 적색의 과피 색소를 가지고 있다. 과실이 가지고 있는
색 및 색소 발현은 유전적인 성질에 의하여 좌우되지만 기상이나 토양 등 환경조건 또는 재
배관리에 의해서도 변화할 수 있다.
녹색색소인 크로로필은 식물의 녹색부분에 있는 엽록체 속의 카로티노이드(황색색소)와 함
께 있다. 이 색소는 광합성의 주역이며 고온이나 광(光)이 많으면 형성된다. 마그네슘이나
질소는 크로로필의 구성요소의 하나이므로 결핍되면 녹색이 소실된다. 철(Fe), 망간(Mn),
구리(Cu) 등이 결핍되어도 녹색은 없어진다. 크로로필은 가을이 되어 저온이 되면 점차 분
해, 소실되고 함께 있던 황색색소가 발현된다. 사과에서는 이러한 상태를 바탕색(地色)이 올
랐다고 표현한다.
황색 색소는 숙기가 되면 나타나는데 이 색소는 카로티노이드계 색소이다. 골든데리셔스 품
종은 크산토필이라고 하는 카로티노이드 색소가 포함되어 있다. 이 색소는 미숙과에도 포함
되어 있지만 녹색 색소인 크로로필에 감추어져 가을이 되어 크로로필이 없어지고 나서야 눈
에 보이게 된다.
적색 색소는 안토시아닌이라고 하는 색소로 과피나 때로는 과육세포에도 포함되어 있는데
당의 축적, 광, 저온에서 발현된다.
안토시아닌은 산성(酸性)에서는 적색으로, 알칼리성에서는 청색으로 발현되는 색소이므로
과실의 산성도, 과실 내 색소분포, 안토시아닌 종류별 함량에 따라 과피색은 미묘하게 변화
한다.
2) 적색 색소의 생성
유전인자 : 동일한 사과라도 다량의 적색 색소를 만들어내는 능력이 있는 스타킹데리셔스
품종과 적색 색소 함량이 극히 적은 골든데리셔스와 같이 안토시아닌의 종류나 색소 발현
정도가 크게 달라지는 것은 품종 고유의 유전적인 특성이다. 적색 색소를 다량으로 생성하
는 능력이 있는 품종은 적색 품종, 능력이 적은 품종은 황색 품종이라고 한다.
당의 축적 잎에서 운반된 당은 여러 : 가지 형태의 물질로 변화해 가는데 PP회로, 쉬키미
산, 페닐피루빈산이라는 물질로 변화한다.
페닐피루빈산으로부터 안토시아닌이 만들어지는 통로와 질소를 이용하여 과실의 비대에 사
용되는 통로가 만들어진다. 따라서 적색을 많이 발현시키기 위해서는 과실 속에 당을 축적
시키고 질소를 억제하여 단백질을 만드는 길을 좁히고 안토시아닌을 만드는 길을 넓힐 필요
가 있다(그림 58, 59).
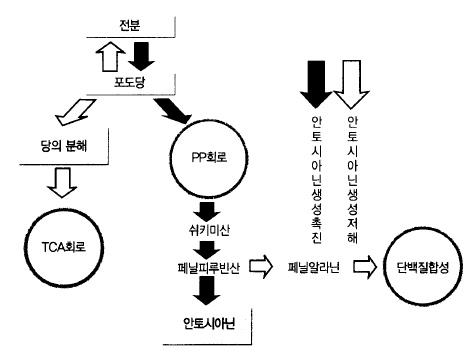
그림 58. 사과 적색색소 안토시아닌의 생성 경로 (Faust, 1965)
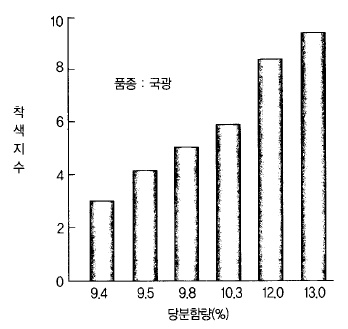
그림 사과 과실의 59. 당분 함량과 착색과의 관계 (일본 아오모리사과시)
광(光) : 식물체 내에 포함되어 있는 안토시아닌 중에는 직접 햇빛이 닿지 않아도 생성되는
것도 있으나 사과의 안토시아닌은 반드시 빛이 닿아야 발현된다. 태양광선에는 파장이 짧고
눈에 보이지 않는 자외선(紫外線), 눈에 보이는 7가지 색의 가시광선(可視光線), 파장이 긴
적외선(赤外線) 등 여러 가지 파장의 광선이 있다. 그 중에 400나노메터(㎚) 전후의 파장을
가진 자외선이 가장 적색을 발현하는 힘이 강하다.
온도(溫度) : 기온이 높은 해는 착색이 지연되고, 선명하지 않다. 욱(매킨토시) 품종을 이용
한 시험결과를 보면 착색의 적온은 15℃이고 20℃이상 고온 또는 10℃이하의 저온에서는
착색이 불량하다. 낮 기온 뿐만 아니라 야간 기온도 안토시아닌 발현에 영향을 미치는데,
기온이 낮은 쪽이 착색을 좋게 한다. 특히 밤 동안은 저온이고 낮 동안 맑은 날씨가 되면
급속히 적색이 발현된다(그림 60). 과실 자체의 온도도 착색에 영향을 미치는데 저온일수록
착색이 빨라진다(그림 61).
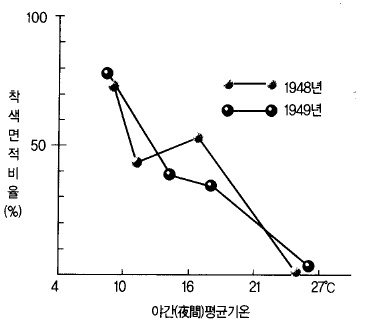
그림 60. 사과의 착색과 야간기온
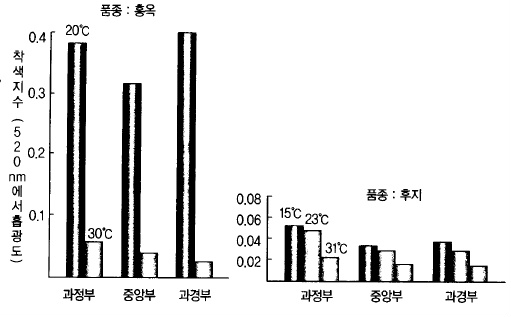
그림 61. 안토시아닌 함량에 미치는 과실 온도의 영향 (苫名, 1980)
고온이 되면 적색이 발현되지 않는 이유는 안토시아닌이 생성되지 않는 반면, 엽록소가 새
로이 만들어지고 분해, 소실이 늦어지기 때문이다.
질소(窒素) 및 수분(水分) : 수체 내에 질소분이나 수분이 적으면 안토시아닌 생성이 많아진
다. 이것은 질소분이 결핍되면 단백질 합성의 길이 좁아져 안토시아닌을 생성하기 위해서
전분이 많이 사용되기 때문이다. 수분이 부족한 경우에는 물에 녹기 어려운 탄수화물이 물
에 녹기 쉬운 형태로 바뀌든지 하여 안토시아닌을 만드는데 유리하게 작용할 것으로 생각된
다(그림 62).

그림 62. 생육시기별 토양의 건조가 사과 생육 및 과실품질에 미치는 영향 (熊代등)
질소나 수분이 많으면 새가지가 번무하여 생장에 탄수화물이 다량으로 사용되든가 햇빛 투
과가 나빠져 잎의 활동이 떨어지기 때문에 착색이 불량해지는 요인으로도 작용한다.
바. 수확전 낙과가 발생한다.
과실이 성숙하고 종자가 노화되면 과실꼭지 기부쪽에 이층(離層)이 형성되어 낙과가 시작된
다(그림 63).
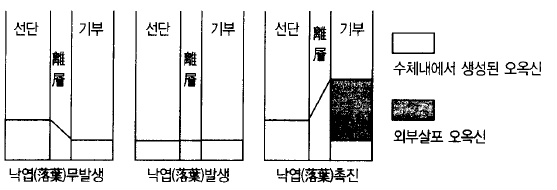
그림 63. 이층(離層)형성 부위에 있어서 오옥신의 구배와 낙과의 관계 (Addicott, 1955)
이러한 낙과 현상을 6월 낙과(조기낙과, June drop)와 대비하여 후기낙과 또는 수확 전 낙
과라고 한다.
이층형성에는 오옥신, 에틸렌, ABA 등의 식물호르몬이 관여하는데 오옥신은 이층형성의 방
지역할을 하고 에틸렌과 ABA는 이층형성을 촉진한다. 과실의 종자에서 생성되고 있던 오옥
신이 종자가 노화되면서 생산이 감소하여 과실쪽의 오옥신 농도가 가지쪽의 오옥신 농도보
다 묽어지면 과실로 양수분의 공급이 불충분하여 에틸렌이 생산되고 ABA가 많아져 이층이
형성된다.
사. 수확기를 알리는 산화(信號)는 여러 가지 있다.
사과의 수확기는 동일한 품종이라도 재배지역, 기후, 재배관리에 따라서 다른데 수확시기를
알리는 신호는 여러 가지 있다. 품종은 유전적인 특성에 따라 만개기로부터 수확까지의 일
수는 거의 결정되어 있다. 그러나 해에 따라서 성숙 정도는 미묘하게 차이가 나므로 전분,
당, 산, 경도 등 내용물의 변화와, 밀발생, 착색정도, 바탕색의 변화, 낙과 정도, 종자의 색
의 변화 등 종합적으로 판단하여야 한다. 특히, 전분, 당, 산, 밀발생 정도, 착색 및 식미가
숙기 판단의 요점이다(그림 64).

그림 64. 요드카리액에 의한 사과 과육의 전분소실정도와 용도별 수확기
(품종 : 후지, 원예연구소, 2003)
5. 휴면기(休眠期)
가. 낙엽이 되면 휴면이 시작된다.
사과나무의 본격적인 휴면은 낙엽과 동시에 시작되는데 나무의 부위에 따라 생장의 늦고 빠
름이 있기 때문에 휴면에 들어가는 시기는 다르다. 특히, 2차 생장한 가지는 휴면에 들어가
는 시기가 상당히 늦어진다. 휴면은 사과나무의 눈(芽)만이 아니고 종자 등 다른 기관에서
도 보이는데 뿌리만은 생장에 적합한 조건만 주어지면 생육을 계속한다.
1) 휴면의 필요성
식물체가 순조로운 생육을 계속하기 위해서는 외계의 악조건으로부터 자기 자신을 어떻게
순응시키고 조절하는가가 중요하다. 추운 겨울에 견디어 나가기 위한 자기 조절이 휴면이
다.
과수의 눈은 인편(鱗片)으로 싸여 있어 빗물의 침입이나 냉기(冷氣)로부터 보호된다. 낙엽된
자리나 과실이 수확된 자리에도 보호조직이 생겨 추위에 대한 준비가 이루어진다. 휴면이
충분히 이루어지지 않은 나무는 눈이 발아, 개학되지 않거나 개화하여도 결실이 불량하다.
2) 자발휴면과 타발휴면
생장에 아무리 적합한 조건이 주어지더라도 생육이 시작되지 않는 휴면을 자발휴면(自發休
眠)이라고 한다. 자발휴면이 완료되기 위해서 사과는 7.2℃ 이하의 온도에서 1,200~1,500
시간 정도 경과하여야 된다. 자발휴면이 종료되면 생장에 적합한 환경이 주어질 때 발아가
시작되는데 실제로는 환경조건이 부적합하기 때문에 여전히 휴면이 계속되는 상태를 타발휴
면(他發休眠)이라고 한다.
우리나라 기상조건에서는 일반적으로 1월 중하순경까지가 자발휴면시기이고 그 이후 3월
하순까지가 타발휴면시기이다. 자발휴면을 타파하는 데에는 저온이 필요하고 반대로 타발휴
면을 타파하기 위해서는 고온이 필요하다.
나. 휴면에도 식물호르몬이 관여한다.
자발휴면이 시작되면 눈이 붙어있는 가지의 전분함량이 증가하고 당은 감소한다. 그리고 휴
면이 가장 깊은 시기에 전분농도가 최고에 이른다. 그 후 감소가 시작되어 자발휴면이 끝날
즈음에는 당이 전분보다 많게 된다(그림 65).
한편 식물호르몬에도 변화가 나타나는데 해가 짧아지면 잎에서 만들어진 생장억제호르몬인
ABA가 증가하여 그것이 눈에 축적되어 휴면에 들어간다. 해가 길어지고 저온을 통과하면
눈 속에는 ABA와의 반대의 작용이 있는 지베렐린이 증가하여 휴면이 타파된다(그림 66).
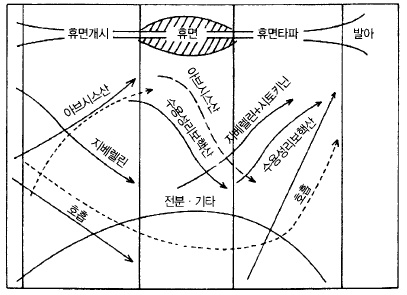
그림 65. 사과나무 휴면에 관계하는 대사작용 (Lavee, 1973)
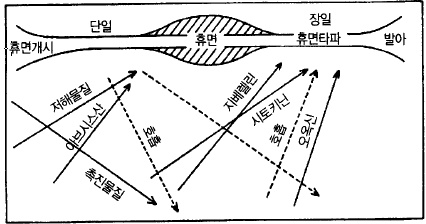
그림 66. 휴면기에 있어서 식물호르몬의 변화(Lavee, 1973)
다. 저온이 내동성(耐凍性)을 증가시킨다.
생육이 계속되고 있는 동안 나무의 내동성은 극히 약하지만 신장이 멎고 가지가 노화되면
내동성은 증가하여 0℃ 이하가 되면 추위에 강하여 진다.
이와 같이 식물체가 저온을 만나면 내동성이 증가하는 것을 하드닝(hardening)이라고 한다.
앞에서 설명한 바와 같이 저온에 의하여 전분이 당으로 변하고 세포액의 농도가 높아지기
때문에 저온에 의한 용액의 동결을 방지하는 힘이 생긴다. 즉, 전분이 당으로 변하여 내동
성을 증가시키기 위해서는 가을까지 수체 내에 충분한 저장양분을 축적하여 두는 것이 중요
하므로 과다착과, 병해충에 의한 잎의 기능 저하, 조기 적엽 등 저장양분을 감소시키는 관
리는 극력 피해야 한다.
라. 수확 후에도 과실은 살아있다.
과실은 나무에 달려 있을 때는 물론 수확 후 저장 중에도 호흡을 하며 생리적인 대사작용을
하고 있다. 그러나 나무에 달려 있을 때와는 달리 외부로부터 양분의 공급이 단절되어 있기
때문에 과실 속에 저정된 양분을 소모하면서 살아가는데 저장양분이 바닥나면 죽음에 이른
다.
저장 중 변화가 현저한 것은 과실수분이 점차 없어지고, 펙틴화합물이 감소하여 과실의 경
도가 떨어진다. 당분은 품종에 따라서 한때는 증가하지만 저장 말기가 되면 감소한다. 감소
폭이 가장 큰 것은 산함량인데 호흡기질로 다량 사용되기 때문이다. 따라서 사과를 오래 살
려두기 위해서는 얼지 않을 정도로 온도를 내리고, 창고 내 공기 중의 가스농도를 조절하여
호흡을 억제시키는 것이 중요하다. 또, 과실에서 수분의 증산을 방지하기 위해서 창고내의
습도를 높여 주는 것이 필요하다(그림 67).
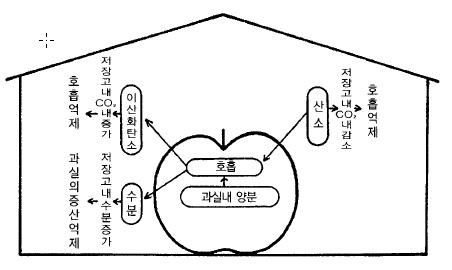
그림 67. 저장고내에서의 과실의 호흡
마 . 사과나무의 지상부와 지하부는 균형을 유지한다.
뿌리에서 흡수된 양수분으로 새가지가 자라고 새 가지 잎에서 만들어진 양분으로 뿌리의 발
육이 이루어진다. 이와 같이 지상부(Top)와 지하부(Root)는 서로 영향을 받으면서 일정한
비율을 유지하는데(그림 68) 이것을 T/R율이라고 한다.
사과나무는 지상부 중량 3에 대하여 지하부는 1의 비율이 보통이지만 수령이나 수세에 따
라서 생육이 달라지므로 ‘지상부 2~5 : 지하부 1’의 비율로 변동한다. 유목기는 지하부보다
지상부가 적어 T/R율이 적지만 수령이 진행됨에 따라 지상부는 목질부가 증가하는데 반해,
뿌리는 오래된 것은 썩어 없어지고 새뿌리로 대체되기 때문에 T/R율은 점차 증가한다.
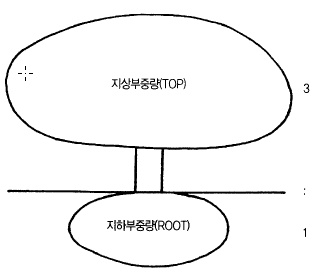
그림 68. 사과나무의 T/R 율
비옥한 토양에서는 양수분이 풍부하므로 뿌리가 길게 자라지 않아도 양수분을 자유로이 흡
수할 수 있기 때문에 뿌리는 세근 이 (細根) 많아지고 뿌리의 중량에 비하여 지상부의 생육이
좋아져 T/R율은 증가한다. 한편, 척박한 토양에서는 뿌리가 양수분을 찾아서 신장하기 때문
에 T/R율은 적어지고, 건조지나 비료를 적게 주는 과수원에서도 같은 경향이다. 이와 같은
토양에서는 관수, 멀칭, 질소비료를 시용함으로써 지상부 발육을 좋게 하여 T/R율을 높일
수 있다.
토양의 좋고 나쁨이나 비배관리의 좋고 나쁨에 따라 T/R율에 변화가 생기는데 휴면기 동안
사과나무의 지상부 상태는 눈에 보이지 않는 뿌리의 생육상태를 보여준다고 할 수 있다.
sa
'식물 자료 > 식물 이야기' 카테고리의 다른 글
| 산기니아(엔젤트럼펫) 키우기 (0) | 2010.05.26 |
|---|---|
| 아라비안자스민 (0) | 2010.05.26 |
| 튜란타(발렌타인쟈스민) 향기나는꽃 키우기 (0) | 2010.05.26 |
| 부룬펠시어 향기나는나무 향기나는꽃 (0) | 2010.05.26 |
| 적하수오키우기, 적하수오묘목구입 (0) | 2010.05.25 |